Biological Coherence in the Built Environment
A New Model of Health: The Built Environment as Physiology

There is a building I have spent time in that does something to me I have never quite been able to name.
It is not beautiful in any obvious way. The proportions are generous. The light moves across the surfaces in a way that changes through the day. There is organic material in it: wood, stone, something soft underfoot. I leave it feeling more settled than when I arrived.
I used to call this atmosphere. I have come to think it is biology.
In medicine we are trained to think about the body as something that happens inside the skin. The environment appears occasionally, as risk factor, as exposure, as context. It has always struck me as an incomplete way of seeing things. We have built a world that the nervous system does not recognise.
That is not a dramatic claim. It is a mechanistic one. And the evidence for it has been building quietly across several disciplines that have not always been talking to each other.
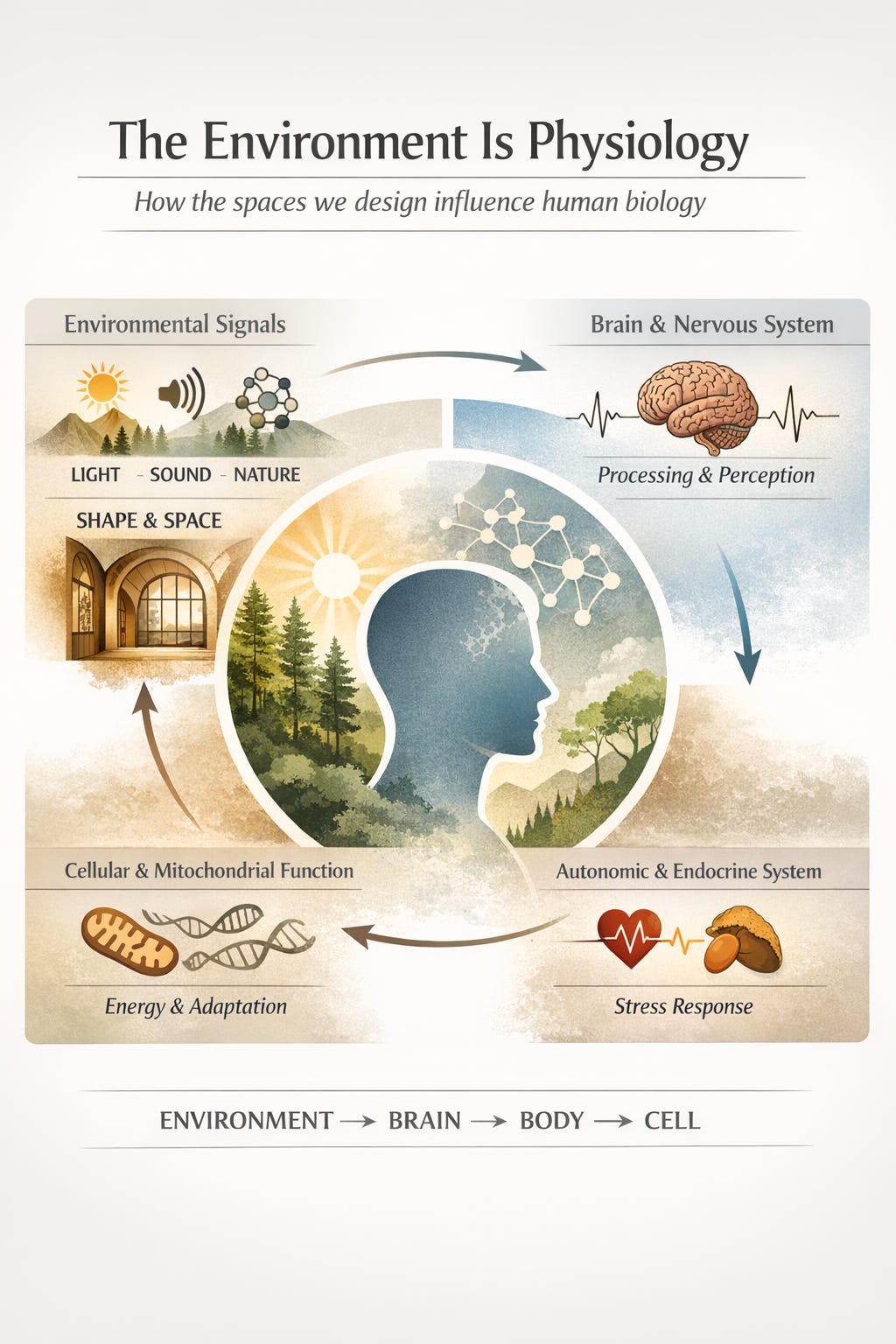
The argument I keep returning to is a simple one, though it sits awkwardly against how we usually talk about health.
The environment is physiology.
We have inherited a model of the body that imagines it as a mostly sealed unit. Something you manage from the inside: sleep better, eat differently, move more, stress less. The environment appears in that model as a backdrop, occasionally significant, mostly peripheral. This framing has always seemed incomplete to me.
The body did not evolve in a sealed container. It evolved in continuous dialogue with its surroundings: light that varied by hour and season, soundscapes that carried information about proximity and safety, visual landscapes of organic complexity and fractal depth. The nervous system, the immune system, the mitochondria, these are not systems that merely tolerate the environment. They read it. Continuously. At a cellular level.
That reading never stops. We simply stopped designing for it.
The question that has been occupying me lately is a specific one: what happens to the biology when the visual environment is chronically incoherent?
We have reasonable evidence about what light does to circadian systems. What sound does to the nervous system. What the absence of microbial diversity does to immune regulation. The visual geometry of built environments has been less mapped from the biological direction. But neuroscience and vision science have been moving toward each other for some time now, and the meeting point is becoming difficult to ignore.
The work of Wilkins, Penacchio, and colleagues offers a useful starting point. The human visual cortex appears to hold a statistical expectation of what the world should look like. Natural scenes have a characteristic structure: organic complexity, fractal-like depth, spatial frequency energy that decreases proportionally as detail increases. When environments match that profile, processing is efficient. When they deviate, too uniform, too repetitive, too flat or aggressively striated, processing demand rises. The mismatch does not register first as an opinion. It registers as a change in cortical activity. The body responds before the mind has formed a view.
From the direction of neuroaesthetics, parallel findings emerge. Curvilinear architectural forms have been shown to engage brain regions associated with reward and positive affect more strongly than rectilinear ones. Spatial geometry, enclosure, ceiling height, the relationship between a person and a volume, shapes emotional appraisal through identifiable neural pathways. Preference, in this context, is not simply cultural. It is biological.
What both lines of work point toward is the same underlying idea: the visual environment is a regulatory input. The geometry of the room you are in is doing something to your nervous system, whether or not you are aware of it.
I have recently been reading more closely, the work of Dr
Cleo Valentine. Dr Valentine and her collaborators at Cambridge have been building what is, in effect, an early mechanistic framework for how architectural environments may accumulate biological load over time. I find this work genuinely important, and still under recognised outside the architecture and neuroscience communities.
Across a series of recent papers, conceptual, physiological, and small-scale experimental, a pattern begins to emerge. Environments that deviate from the statistical structure of natural scenes appear to increase processing demand in the visual system, with downstream effects on autonomic regulation. In some studies this shows up as shifts in heart rate variability. In others, as changes in cortical activity measured by EEG. The individual signals are modest. The samples are small. But the direction is coherent.
One strand of this work frames the process as a form of architectural allostatic load: sustained, low-grade activation of stress pathways driven not by discrete events, but by the background conditions of the environment itself. The body does not distinguish particularly well between sources of stress input. Repeated activation, whatever the origin, feeds into the same neuroendocrine systems.
There are early indications of what that might mean in practice. Curved and proportionate architectural forms have been associated with lower physiological stress markers compared to more angular or visually irregular spaces. In pilot EEG work, environments with higher biophilic content have been linked to shifts in brain activity consistent with reduced processing strain, though the interpretation remains indirect and requires further validation.
More recent studies combining autonomic and cortical measures add another layer. The cortical signal is not yet consistently detectable at this scale. But the autonomic system appears to respond reliably. The body is reacting to what the visual system is processing, even when conscious awareness remains unchanged.
What is emerging is not a finished model, but a plausible chain. Visual environment shapes cortical demand. Cortical demand influences autonomic regulation. Repeated over time, those signals enter the neuroendocrine system. And from there, they reach the cell.
From the biological direction, the intracellular layer is where this becomes harder to ignore.
The HPA axis does not terminate at cortisol. Cortisol reaches the mitochondria. Mitochondria are sensitive to the organism’s perceived threat state: they read the neuroendocrine environment and adjust energy production accordingly. Under sustained threat signalling, they shift from generative to defensive modes. ATP output narrows. BDNF, the growth factor that supports neurogenesis and synaptic plasticity, is partly dependent on mitochondrial capacity. When that capacity is constrained, the hippocampus does not receive the metabolic support it requires.
The subjective experience of this is rarely illness. It is quieter than that. A subtle erosion of capacity. The tiredness that sleep does not fully resolve. The cognitive effort that sits slightly higher than it should. These are experiences people tend to attribute entirely to how they are living. And often that attribution is correct. But the environments in which those lives are lived are part of the same equation, and have almost never been measured.
Running alongside this is a second pathway. The retinal cells that set the body’s central clock are reading light continuously for intensity, spectrum, and timing. Buildings that flatten or fragment natural daylight disrupt circadian entrainment across multiple systems. Circadian disruption alters NAD⁺ metabolism, with downstream effects on mitochondrial function and DNA repair. Geometry and light are not separate considerations. They converge on the same biology. Light is not only a visual signal but a metabolic one, interacting directly with mitochondrial function through wavelength-specific pathways that are only beginning to be fully understood.
A recent randomised crossover study offers a grounding note. Participants performing a cognitively demanding task in the presence of a living wall showed higher parasympathetic tone than those exposed to a shelf of objects. The effect was present in both real and virtual environments, but only the real environments shifted subjective affect.
The body registers the difference between organic material and its representation.
It is important to be precise about what the evidence does and does not yet support.
The mechanistic framework is coherent. The individual studies are often small, many are early-stage, and the full causal chain across long timescales in humans remains to be established. That work is ahead of us.
But the direction is consistent. Across vision science, neuroaesthetics, stress physiology, chronobiology, and emerging architectural neuroscience, the same signal appears: the environments we build are not passive. They are active participants in physiological regulation.
We have simply not been treating them as such.
We spend most of our lives indoors. Those interiors have been designed with remarkable precision for cost, efficiency, and convention, and almost none for the biological systems that inhabit them.
What is beginning to emerge, slowly, and across disciplines, is a different kind of design question. One that treats the geometry of a room as a health input. The curvature of a surface as a regulatory variable. The presence of organic material as a signal to a nervous system that evolved in its presence.
The medicine the body runs is not separate from the container it runs in.
We have been building containers as if it were.
Wilkins AJ, Penacchio O, Leonards U. The built environment and its patterns. SDAR Journal. 2018.
Coburn A, Vartanian O, Chatterjee A, et al. Psychological and neural responses to architectural interiors. Cortex. 2020;126:217–241.
Shemesh A, Talmon R, Karp O, et al. A neurocognitive study of the emotional impact of geometrical criteria of architectural space. Architectural Science Review. 2021;64(4):394–407.
Valentine C. Architectural allostatic overloading: A new framework for understanding stress in the built environment. 2023.
Valentine C. Impact of architectural form on physiological stress: A systematic review. 2024.
Valentine C, Steffert T, Mitcheltree H, Steemers K. Architectural neuroimmunology: A pilot study. Buildings. 2024;14(5):1292.
Valentine C, Mitcheltree H, Sjövall IAK, Khalil MH. Architecturally mediated allostasis and neurosustainability. Brain Sciences. 2025.
Serra H, et al. Biophilic interventions in real and virtual environments reduce stress during cognitively demanding tasks. Scientific Reports. 2025;15:39642.
Valentine C, Hosking I, Wilkins AJ, Mitcheltree H, Smith C, Butters E, Penacchio O. Impact of architecture façade design on neurophysiological stress using fNIRS and HRV. Buildings. 2026.
I really appreciate this perspective, especially the idea that biological systems lose flexibility before disease appears. What I keep asking, though, is where coherence actually breaks. Because from a physiological standpoint, coherence is not just a state, it is a function of transport. A system can appear coherent or incoherent depending on whether energy actually reaches the cell. In that sense, what we call coherence might simply be the visible outcome of a stable gradient, and what we call decoherence a failure of transport across interfaces.